Voltar Clínica Antienvelhecimento Quiropraxia e Equipe de Medicina Funcional. Nosso corpo está em uma batalha constante e sem fim pela sobrevivência. As células nascem, as células são destruídas. Os cientistas estimam que cada célula deve resistir a mais de 10,000 ataques individuais de espécies reativas de oxigênio (ROS) ou radicais livres. Sem falha, o corpo tem um incrível sistema de autocura que resiste ao ataque e reconstrói o que foi danificado ou destruído. Esta é a beleza do nosso design.
Compreender a biologia do envelhecimento e traduzir o conhecimento científico em intervenções que melhoram a saúde na velhice por meio de tratamentos. É útil ter uma visão clara e consensual sobre o que exatamente constitui um tratamento anti-envelhecimento.
Desde antes da busca de Ponce de Leon pela longevidade, o homem sempre foi seduzido pela chance da juventude eterna. A quiropraxia com seu movimento de saúde é um método poderoso de estabilizar e aumentar essa capacidade de autocura. O Dr. Alex Jimenez discute os conceitos que cercam a pandora anti-envelhecimento.
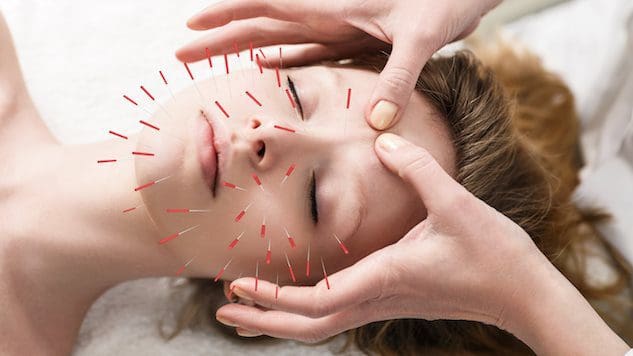
Para indivíduos que desejam melhorar ou manter a saúde da pele, a incorporação da acupuntura pode ajudar a melhorar a pele e combater o processo de envelhecimento?
Acupuntura Cosmética
A acupuntura cosmética segue a prática tradicional da acupuntura de inserção de agulhas. O objetivo é reverter os sinais de envelhecimento e melhorar a saúde da pele. Às vezes é chamado de rejuvenescimento facial por acupuntura, que tem sido usado como uma alternativa aos liftings cirúrgicos e outros procedimentos convencionais. Estudos preliminares examinaram como ele pode ajudar a remover manchas da idade, levantar pálpebras caídas e reduzir rugas. (Younghee Yun et al., 2013)
Como funciona a acupuntura
Na medicina tradicional chinesa ou MTC, a acupuntura tem sido usada há muito tempo para melhorar o fluxo de energia – qi ou chi – por todo o corpo. Acredita-se que essa energia circule por vias energéticas conhecidas como meridianos. Quando ocorrem problemas de saúde, segundo a MTC, ocorrem obstruções ou bloqueios na circulação.
Os acupunturistas podem restaurar a circulação/fluxo ideal e melhorar a saúde inserindo agulhas em pontos de acupuntura específicos. (Institutos Nacionais de Saúde, 2007)
Acupuntura Cosmética
Diz-se que a acupuntura cosmética melhora a saúde da pele e atua como um tratamento anti-envelhecimento, estimulando a produção de colágeno. Esta proteína é um componente importante da pele. A camada interna da pele perde colágeno e firmeza à medida que o corpo envelhece. No entanto, são necessárias mais pesquisas para apoiar a afirmação de que a acupuntura pode promover a produção de colágeno. Alguns sugerem que a acupuntura cosmética ajuda a rejuvenescer a pele, melhorando a energia geral do corpo. Um estudo descobriu que os indivíduos observaram melhorias após cinco sessões de acupuntura cosmética facial. (Younghee Yun et al., 2013) No entanto, recomenda-se que dez tratamentos sejam realizados uma ou duas vezes por semana para obter melhores resultados. Depois disso, os tratamentos de manutenção são feitos a cada quatro a oito semanas. Ao contrário do Botox ou dos preenchimentos dérmicos, a acupuntura cosmética não é uma solução rápida. O foco é criar mudanças de longo prazo na pele e no corpo, o que significa melhorar:
Quando as agulhas são inseridas na pele, criam feridas conhecidas como microtraumas positivos. As habilidades naturais de cura e reparação do corpo são ativadas quando ele sente essas feridas. Essas punções estimulam os sistemas linfático e circulatório, que levam nutrientes e oxigênio às células da pele, nutrindo-as de dentro para fora.
Isso ajuda a uniformizar a tez e promove o brilho da pele.
Os microtraumas positivos também estimulam a produção de colágeno.
Isso ajuda a melhorar a elasticidade, minimizando linhas e rugas.
Alternativas
Vários remédios naturais podem ajudar a melhorar a saúde da pele e oferecer benefícios antienvelhecimento. As ceramidas são uma molécula de gordura encontrada naturalmente na camada superior da pele e um ingrediente usado em produtos para a pele. Estes podem proteger contra o ressecamento da pele relacionado ao envelhecimento. (L Di Marzio 2008) Pesquisas preliminares sugerem que a aplicação de chá branco na pele pode combater a degradação do colágeno e da elastina – uma proteína que apoia a elasticidade da pele e previne a flacidez). Também há evidências de que substâncias naturais como óleo de argan, óleo de borragem e espinheiro podem oferecer benefícios hidratantes que podem melhorar a pele.(Tamsyn SA Thring et al., 2009)
Embora sejam necessárias mais evidências da acupuntura cosmética, a integração da acupuntura pode ajudar a controlar o estresse e melhorar a saúde geral. Indivíduos que consideram a acupuntura cosmética devem consultar seu médico primário para ver se ela é adequada para eles.
Melhorando a saúde juntos: adotando avaliação e tratamento multidisciplinar
Referências
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS e Choi, I. (2013). Efeito da acupuntura cosmética facial na elasticidade facial: um estudo piloto aberto e de braço único. Medicina complementar e alternativa baseada em evidências: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
O Centro Nacional de Medicina Complementar e Alternativa. (2007). Acupuntura: uma introdução. Site do Centro Nacional de Medicina Complementar e Alternativa. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH e Tsuji, R. (2021). Confiabilidade e Validade da Folha de Verificação Facial (FCS): Lista de Verificação para Auto-Satisfação com Acupuntura Cosmética. Medicamentos (Basileia, Suíça), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008). Aumento dos níveis de ceramida na pele em indivíduos idosos após uma aplicação tópica de curto prazo de esfingomielinase bacteriana de Streptococcus thermophilus. Jornal internacional de imunopatologia e farmacologia, 21(1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P. e Naughton, DP (2009). Atividades anticolagenase, antielastase e antioxidante de extratos de 21 plantas. Medicina complementar e alternativa BMC, 9, 27. doi.org/10.1186/1472-6882-9-27
Manter a coluna vertebral de um indivíduo em sua melhor forma significa menos dor e mais mobilidade, flexibilidade e liberdade. O corpo se desgasta e é um efeito natural do envelhecimento que acontece com cada um de nós. Problemas na coluna vertebral relacionados ao envelhecimento podem se tornar sérios se não forem tratados e implementados com exercícios, alongamento e manutenção quiroprática.
Envelhecimento e as costas
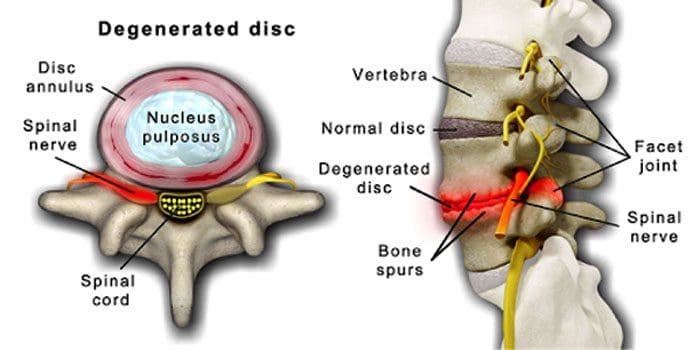
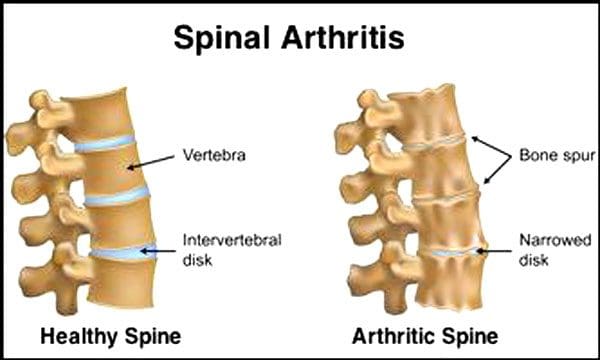
É normal que os discos espinhais e as articulações se deteriorem com a idade. A estenose espinhal ou o estreitamento do canal vertebral também podem fazer parte do processo de envelhecimento. Duas condições causadas pelo envelhecimento são doença degenerativa do disco e artrite que também pode incluir enrijecimento dos ligamentos espinhais e osteoporose.
A doença degenerativa do disco é experimentada por 40% dos indivíduos com 40 anos de idade
Aumenta para 80% para indivíduos com 80 anos de idade ou mais.
Centra-se em torno de discos que mudam gradualmente de principalmente água para principalmente gordura.
Quando é gordo, os discos se estreitam e perdem elasticidade.
Os Centros de Controle e Prevenção de Doenças afirmam que 23% dos adultos americanos têm artrite. Esta é uma condição que afeta principalmente as articulações. As articulações ficam inchadas, o que reduz a amplitude de movimento e pode afetar os nervos espinhais, causando dor, fraqueza e ciática. Com o tempo, os ligamentos ao redor e na coluna se enrijecem, reduzindo a amplitude de movimento, causando estenose. A perda óssea, ou osteoporose, é causada por mudanças nos hormônios e outros fatores, como nutrição. O envelhecimento é um processo natural, mas os indivíduos podem ajudar a manter a sua coluna vertebral em sua melhor forma, independentemente da idade.
Praticando uma postura saudável
Logo de cara mecânica corporal saudável adequada é uma obrigação. Ficar atento e atento à postura corporal mantém o alinhamento e o equilíbrio do corpo. A postura saudável ajudará a reduzir os efeitos de:
Estenose espinal
Doença degenerativa do disco
Herniação
Risco de fraturas da coluna vertebral
Praticar a postura adequada inclui:
Reduzir desleixo
Certifique-se de que a estação de trabalho está em sua melhor forma e ergonomicamente sólida
Certifique-se de dobrar os joelhos ao levantar e manter a coluna o mais vertical possível.
Ioga
Ioga pode ser altamente benéfico para uma coluna mais saudável e jovem. A ioga preenche três áreas para manter a coluna vertebral em sua melhor forma. Isso inclui:
Exercício regular
Mantém flexibilidade
Alcança o peso corporal ideal
Yoga é uma atividade anti-envelhecimento para a coluna vertebral. Porque isso:
Mantém a força
Flexibilidade
Postura
Equilíbrio
Pode ser útil para uma variedade de doenças da coluna vertebral, especificamente dor de artrite
As quedas podem causar ferimentos graves. A ioga também pode ajudar a trabalhar o equilíbrio.
Consulte um quiroprático
A medicina preventiva é a chave para manter o corpo saudável, jovem e o mais forte possível. Um exame de Quiropraxia pode determinar se há algum problema de coluna e um diagnóstico para desenvolver um plano de tratamento ideal. Se a função corporal é limitada por causa da dor nas costas e / ou pernas, entre em contato com a Clínica de Quiropraxia Médica de Lesões e a Clínica de Medicina Funcional e coloque a coluna vertebral de volta em sua melhor forma.
Composição do corpo
Exercício / Estabilidade Bola Curls
Esse exercício trabalha grupos musculares específicos para a força da coluna e inclui:
Isquiotibiais
Glutes
Abdominais profundos
Abdutores e rotadores do quadril
Exercícios como este são uma das maneiras mais eficazes de construir força funcional e resistência nos isquiotibiais, quadris e prevenir lesões. Para fazer este treino:
Deite-se de costas com os joelhos dobrados
Levante as pernas para que a planta dos pés fique em cima de uma bola de exercícios
Role suas pernas até que fiquem retas
Mantenha a posição por um ou dois segundos
Retorne ao topo do movimento enquanto aperta os tendões
Trabalhar esses músculos ajudará a tornar os movimentos de agachamento, estocadas ou flexões mais fáceis para a coluna.
Isenção de responsabilidade da postagem do blog do Dr. Alex Jimenez
O escopo de nossas informações se limita a artigos, tópicos e discussões sobre quiropraxia, musculoesquelética, medicamentos físicos, bem-estar e questões delicadas de saúde e / ou medicina funcional. Usamos protocolos funcionais de saúde e bem-estar para tratar e apoiar o tratamento de lesões ou distúrbios do sistema músculo-esquelético. Nossas postagens, tópicos, assuntos e percepções cobrem questões clínicas, questões e tópicos que se relacionam e apoiam direta ou indiretamente nosso escopo clínico de prática. *
Nosso escritório fez uma tentativa razoável de fornecer citações de apoio e identificou o estudo de pesquisa relevante ou estudos que apoiam nossas postagens. Também disponibilizamos cópias de estudos de pesquisa de apoio ao conselho e ou ao público mediante solicitação. Entendemos que cobrimos questões que requerem uma explicação adicional sobre como isso pode ajudar em um plano de cuidados ou protocolo de tratamento específico; portanto, para discutir o assunto acima, sinta-se à vontade para perguntar ao Dr. Alex Jimenez ou entre em contato conosco pelo telefone 915-850-0900. O (s) provedor (es) licenciado (s) no Texas e Novo México *
Os alimentos que ingerimos podem ser benéficos ou prejudiciais à nossa saúde. A má nutrição pode causar uma variedade de problemas de saúde, incluindo obesidade, doenças cardiovasculares e diabetes tipo 2. Enquanto isso, a nutrição adequada pode fazer você se sentir energizado, reduzir o risco de problemas de saúde e ajudar a manter e regular um peso saudável. Se você deseja promover a longevidade, precisa alimentar seu corpo com bons alimentos. No artigo a seguir, listaremos vários bons alimentos que podem ajudar a promover a longevidade, ajudando também a melhorar a saúde e o bem-estar em geral.
Vegetais crucíferos
Os vegetais crucíferos têm a capacidade única de alterar nossos hormônios, ativar o sistema de desintoxicação natural do corpo e até mesmo reduzir o crescimento de células cancerosas. Devem ser mastigados bem ou comidos desfiados, picados, espremidos ou misturados para liberar suas propriedades benéficas. O sulforafano, encontrado em vegetais crucíferos, também ajuda a proteger a parede dos vasos sanguíneos da inflamação que pode causar doenças cardíacas. Vegetais crucíferos, como couve, repolho, couve de Bruxelas, couve-flor e brócolis são vários dos alimentos mais ricos em nutrientes do mundo.
Salada Verde
Os vegetais folhosos crus têm menos de 100 calorias por libra, o que os torna o alimento perfeito para perda de peso. Comer mais saladas também foi associado ao risco reduzido de ataque cardíaco, derrame, diabetes e vários tipos de câncer. Os vegetais folhosos crus também são ricos em folato essencial da vitamina B, além de luteína e zeaxantina, carotenóides que podem ajudar a proteger os olhos. Fitoquímicos lipossolúveis, como carotenóides, encontrados em saladas como alface, espinafre, couve, couve e mostarda também têm efeitos antioxidantes e anti-inflamatórios no organismo.
Nozes
As nozes são um alimento com baixo índice glicêmico e uma ótima fonte de gorduras saudáveis, proteínas vegetais, fibras, antioxidantes, fitoesteróis e minerais, o que também ajuda a reduzir a carga glicêmica de uma refeição inteira, tornando-a parte essencial de um anti-diabetes dieta. Independentemente de sua densidade calórica, comer nozes pode ajudar a promover a perda de peso. As nozes também podem reduzir o colesterol e ajudar a reduzir o risco de doença cardíaca.
Tratamentos para
As sementes, assim como as nozes, também fornecem gorduras saudáveis, antioxidantes e minerais; no entanto, elas têm mais proteínas e são ricas em minerais. Sementes de chia, linho e cânhamo são ricas em gorduras ômega-3. As sementes de chia, linho e gergelim também são lignanas ricas ou fitoestrógenos que combatem o câncer de mama. Além disso, as sementes de gergelim são ricas em cálcio e vitamina E, e as sementes de abóbora são ricas em zinco.
Berries
Bagas são frutas ricas em antioxidantes que podem ajudar a promover a saúde do coração. Estudos de pesquisa em que os participantes comeram morangos ou mirtilos diariamente por várias semanas relataram melhorias na pressão sanguínea, colesterol total e LDL e até sinais de estresse oxidativo. As bagas também têm propriedades anti-câncer e demonstraram ajudar a prevenir o declínio cognitivo associado ao envelhecimento.
Romã
O fitoquímico mais conhecido nas romãs, a punicalagina, é responsável por mais da metade da atividade antioxidante da fruta. Os fitoquímicos da romã têm benefícios anticancerígenos, cardioprotetores e saudáveis para o cérebro. Em um estudo, os idosos que ingeriram suco de romã diariamente por 28 dias tiveram um desempenho melhor em um teste de memória em comparação com aqueles que ingeriram uma bebida placebo.
Feijões
Comer feijão e outras leguminosas pode ajudar a equilibrar o açúcar no sangue, reduzir o apetite e proteger contra o câncer de cólon. O feijão é um alimento anti-diabetes que pode ajudar a promover a perda de peso porque é digerido lentamente, o que diminui o aumento de açúcar no sangue após uma refeição e ajuda a prevenir o desejo por comida, promovendo a saciedade. Verificou-se que comer feijão e outras leguminosas duas vezes por semana diminui o risco de câncer de cólon. Comer feijão e outras leguminosas, como feijão vermelho, feijão preto, grão de bico, lentilha e ervilha, também fornece proteção significativa contra outros tipos de câncer.
Cogumelos
Comer cogumelos regularmente está associado a um risco reduzido de câncer de mama. Os cogumelos branco e Portobello são especialmente benéficos contra o câncer de mama, porque possuem inibidores da aromatase ou compostos que inibem a produção de estrogênio. Os cogumelos demonstraram ter efeitos anti-inflamatórios, além de proporcionar atividade celular imune aprimorada, prevenção de danos ao DNA, crescimento lento das células cancerígenas e inibição da angiogênese. Os cogumelos sempre devem ser cozidos, pois os cogumelos crus têm um produto químico potencialmente cancerígeno conhecido como agaritina, que é reduzido significativamente pelo cozimento.
Cebolas e Alho
Cebola e alho fornecem benefícios cardiovasculares e do sistema imunológico, além de efeitos antidiabéticos e anticâncer. Estes também foram associados a um menor risco de câncer de estômago e próstata. Cebola e alho são conhecidos por seus compostos organossulfurados, que ajudam a impedir o desenvolvimento de cânceres por desintoxicantes substâncias cancerígenas, diminuindo o crescimento de células cancerígenas e bloqueando a angiogênese. Cebola e alho também têm altas concentrações de antioxidantes flavonóides promotores de saúde, que têm efeitos anti-inflamatórios que podem ajudar a fornecer prevenção ao câncer.
Tomates
Os tomates são ricos em uma variedade de nutrientes, como licopeno, vitamina C e E, beta-caroteno e antioxidantes flavonóides. O licopeno pode ajudar a proteger contra o câncer de próstata, danos à pele por UV e? doença cardiovascular. O licopeno é melhor absorvido quando os tomates são cozidos. Uma xícara de molho de tomate tem cerca de 10 vezes a quantidade de licopeno que uma xícara de tomate cru picado. Lembre-se também de que os carotenóides, como o licopeno, são mais bem absorvidos quando acompanhados de gorduras saudáveis, portanto, saboreie seus tomates em uma salada com nozes ou molho à base de nozes para obter benefícios nutricionais extras.
Os alimentos que ingerimos podem ser benéficos ou prejudiciais à nossa saúde. A má nutrição pode causar uma variedade de problemas de saúde, incluindo obesidade, doenças cardiovasculares e diabetes tipo 2. Enquanto isso, a nutrição adequada pode fazer você se sentir energizado, reduzir o risco de problemas de saúde e ajudar a manter e regular um peso saudável. Se você deseja promover a longevidade, precisa alimentar seu corpo com bons alimentos. Bons alimentos também podem ajudar a reduzir a inflamação associada a uma variedade de problemas de saúde, incluindo dores nas articulações e artrite. Profissionais de saúde, como quiropráticos, podem oferecer conselhos sobre dieta e estilo de vida para ajudar a promover a saúde e o bem-estar. No artigo a seguir, listaremos vários bons alimentos que podem ajudar a promover a longevidade. - Dr. Alex Jimenez DC, CCST Insight
Suco de beterraba picante
Dose: 1 Tempo de cozimento: 5-10 minutos
1 toranja, descascada e fatiada
1 maçã, lavada e cortada
1 beterraba inteira e folhas, se as tiver, lavadas e fatiadas
Botão de gengibre de 1 polegada, enxaguado, descascado e picado
Suco de todos os ingredientes em um espremedor de alta qualidade. Melhor servido imediatamente.
Apenas uma cenoura fornece toda a sua ingestão diária de vitamina A
Sim, comer apenas uma cenoura fervida de 80g (2 oz) fornece beta-caroteno suficiente para que seu corpo produza 1,480 microgramas (mcg) de vitamina A (necessária para a renovação das células da pele). Isso é mais do que a ingestão diária recomendada de vitamina A nos Estados Unidos, que é cerca de 900mcg. É melhor comer cenouras cozidas, pois isso amolece as paredes celulares, permitindo que mais beta-caroteno seja absorvido. Adicionar alimentos mais saudáveis à dieta é uma ótima maneira de melhorar sua saúde geral.
O escopo de nossas informações é limitado a artigos, tópicos e discussões sobre quiropraxia, musculoesquelética, medicamentos físicos, bem-estar e questões delicadas de saúde e / ou medicina funcional. Usamos protocolos funcionais de saúde e bem-estar para tratar e apoiar o tratamento de lesões ou distúrbios do sistema músculo-esquelético. Nossas postagens, tópicos, assuntos e percepções cobrem questões clínicas, questões e tópicos que se relacionam e apoiam direta ou indiretamente nosso escopo clínico de prática. * Nosso escritório fez uma tentativa razoável de fornecer citações de apoio e identificou o estudo de pesquisa relevante ou estudos que apóiam nossas postagens. Também disponibilizamos cópias de estudos de pesquisa de apoio ao conselho e ou ao público mediante solicitação. Entendemos que cobrimos questões que requerem uma explicação adicional sobre como isso pode ajudar em um plano de cuidados ou protocolo de tratamento específico; portanto, para continuar a discutir o assunto acima, sinta-se à vontade para perguntar ao Dr. Alex Jimenez ou entre em contato conosco em 915-850-0900. O (s) provedor (es) licenciado (s) no Texas * e Novo México *
Curadoria do Dr. Alex Jimenez DC, CCST
Referências:
Joel Fuhrman, MD. 10 melhores alimentos que você pode comer para viver mais e se manter saudável. Verywell Health, 6 de junho de 2020, www.verywellhealth.com/best-foods-for-longevity-4005852.
Dowden, Angela. O café é uma fruta e outros fatos alimentares incrivelmente verdadeiros. MSN Estilo de vida, 4 de junho de 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Pele avermelhada, especialmente nas palmas das mãos?
Pele ou cabelos secos ou escamosos?
Acne ou pele doentia?
Unhas fracas?
Edema?
Se você estiver enfrentando alguma dessas situações, seus peptídeos de colágeno podem estar baixos.
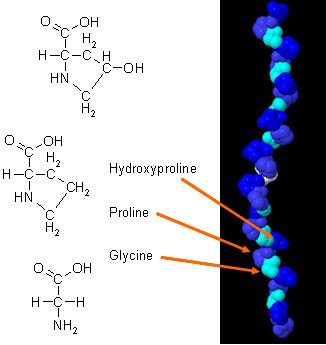
Há foram novos estudos sobre como o colágeno pode melhorar a composição corporal quando combinado com exercícios diários. O colágeno no corpo tem uma composição única de aminoácidos que desempenha um papel essencial na anatomia do corpo. A proteína de colágeno é uma fonte concentrada de glicina, prolina e hidroxiprolina, e quando está sendo comparada a todas as outras proteínas da dieta, torna o colágeno uma possível opção prática como proteína estrutural.
In um estudo 2015, os pesquisadores demonstraram como suplementos de colágeno eficientes podem melhorar a composição corporal em homens ativos. Os resultados mostram como cada indivíduo do sexo masculino está participando do treinamento com pesos pelo menos três vezes por semana e precisa suplementar com pelo menos 15 gramas de peptídeos de colágeno para obter o máximo de saúde. As avaliações fornecidas pelo teste são: teste de força, análise de bioimpedância (BIA) e biópsias musculares. Esses testes garantem que os indivíduos do sexo masculino tenham um bom desempenho após tomar os suplementos de colágeno, e os resultados mostram como sua massa corporal aumentou a massa corporal livre de gordura. Outro estudo mostrou como a suplementação de proteína de colágeno, quando combinada com o treinamento de resistência, pode aumentar a massa muscular e a força muscular tanto em idosos quanto em pessoas com sarcopenia.
Propriedades benéficas com colágeno
Tem muitas propriedades benéficas que os suplementos de colágeno podem fornecer ao corpo quando é consumido. Existem colágeno e gelatina hidrolisados e podem ajudar a melhorar a estrutura da pele de uma pessoa. Embora não existam muitos estudos sobre suplementos de colágeno, existem excelentes promessas para as áreas do corpo. Eles são:
Massa muscular: Os suplementos de colágeno, quando combinados ao treinamento de força, podem aumentar a massa muscular e a força do corpo.
Artrite: Os suplementos de colágeno podem ajudar as pessoas com osteoartrite. Estudos mostram que quando as pessoas com osteoartrite tomam suplementos de colágeno, descobriram um declínio maciço na dor que estavam sentindo.
Elasticidade da pele: Dentro um estudo 2014, afirmou que mulheres que tomaram suplementos de colágeno e mostraram melhorias na elasticidade da pele. O colágeno também pode ser usado em tratamentos tópicos para ajudar a melhorar a aparência da pele de uma pessoa, minimizando linhas finas e rugas.
Não apenas os suplementos de colágeno fornecem propriedades benéficas para as áreas específicas do corpo, mas existem os quatro principais tipos de colágeno e quais são seus papéis no corpo humano, bem como suas funções:
Tipo 1: O colágeno tipo 1 levou em conta 90% do colágeno do corpo e é composto de fibras densamente compactadas que fornecem estruturas para a pele, ossos, tecidos conjuntivos e dentes que estão no corpo.
Tipo 2: O colágeno tipo 2 é constituído por fibras frouxamente encontradas na cartilagem elástica, o que ajuda a amortecer as articulações do corpo.
Tipo 3: O colágeno tipo 3 ajuda a apoiar a estrutura dos músculos, órgãos e artérias que garantem que o corpo esteja funcionando corretamente.
Tipo 4: O colágeno tipo 4 é encontrado nas camadas da pele de todas as pessoas e ajuda na filtração do corpo.
Como esses quatro tipos de colágeno estão no corpo, é essencial saber que o colágeno pode diminuir naturalmente ao longo do tempo com a idade, pois o corpo produz uma menor qualidade menor de colágeno. Um dos sinais visíveis da diminuição do colágeno é quando a pele do corpo humano se torna menos firme e flexível, além de enfraquecer a cartilagem devido ao envelhecimento.
Fatores que podem danificar o colágeno
Embora o colágeno possa diminuir naturalmente com a idade, muitos fatores podem destruir os colágenos que são prejudiciais à pele. Os fatores prejudiciais podem incluir:
Açúcar e Carboidratos: Açúcares refinados e carboidratos pode interferir com a capacidade do colágeno de se auto-regenerar na pele. Portanto, ao minimizar o consumo de açúcar e carboidratos no corpo, ele pode reduzir os efeitos da disfunção vascular, renal e do tecido cutâneo.
Exposição ao sol: Embora obter sol suficiente possa ajudar uma pessoa a aproveitar o dia, no entanto, ficar exposto ao sol por um longo período pode causar danos para a pele e destruir peptídeos de colágeno. Os efeitos da superexposição do sol podem fazer com que a pele envelheça e produza estresse oxidativo no corpo.
Fumar: Quando uma pessoa fuma, pode reduzir a produção de colágeno no corpo, causando rugas prematuras, e se o corpo for ferido, o processo de cicatrização será mais lento e pode levar a doenças no corpo.
Doenças autoimunes: Algumas doenças autoimunes também podem prejudicar a produção de colágeno, como o lúpus.
Conclusão
O colágeno é vital para o corpo, pois ajuda a pele a ser suave e firme. Naturalmente, ele diminui à medida que a pessoa envelhece, portanto, tomar suplementos de colágeno pode garantir que o corpo possa funcionar corretamente. Quando fatores prejudiciais estão afetando o corpo, eles podem parar ou até danificar a produção de colágeno e acelerar o processo de formação de rugas prematuras, fazendo uma pessoa parecer mais velha do que é. Alguns Produtos pode ajudar a atividade celular do corpo, fornecendo estabilidade, biodisponibilidade e conforto digestivo mais excelentes.
O escopo de nossas informações limita-se a problemas de saúde quiroprática, músculo-esquelética e nervosa ou a artigos, tópicos e discussões sobre medicina funcional. Utilizamos protocolos funcionais de saúde para tratar lesões ou distúrbios do sistema músculo-esquelético. Nosso escritório fez uma tentativa razoável de fornecer citações de apoio e identificou o (s) estudo (s) de pesquisa relevante (s) que apóiam nossos posts. Também disponibilizamos cópias dos estudos de apoio à diretoria e / ou ao público, mediante solicitação. Para discutir melhor o assunto acima, não hesite em perguntar ao Dr. Alex Jimenez ou entre em contato em 915-850-0900.
Referências:
Bosch, Ricardo, et al. Mecanismos de Fotoenvelhecimento e Fotocarcinogênese Cutânea e Estratégias Fotoprotetoras com Fitoquímicos. Antioxidantes (Basel, Suíça), MDPI, 26 de março de 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Danby, F. William. Nutrição e envelhecimento da pele: açúcar e glicação. Clínicas em Dermatologia, Biblioteca Nacional de Medicina dos EUA, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Jennings, Kerri-Ann. Colágeno - O que é e para que serve? Healthline, 9 de setembro de 2016, www.healthline.com/nutrition/collagen.
Jurgelewicz, Michael. Novo estudo demonstra os benefícios dos peptídeos de colágeno para melhorar a composição corporal combinada com exercícios. Projetos para a Saúde, 31 de maio de 2019, blog.designsforhealth.com/node/1031.
Knuutinen, A, et al. Fumo afeta a síntese de colágeno e a renovação da matriz extracelular na pele humana. O Jornal Britânico de Dermatologia, Biblioteca Nacional de Medicina dos EUA, abril 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E, et al. A suplementação oral de peptídeos de colágeno específicos tem efeitos benéficos na fisiologia da pele humana: um estudo duplo-cego controlado por placebo. Farmacologia e Fisiologia da Pele, Biblioteca Nacional de Medicina dos EUA, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G, et al. Efeito do novo extrato hidrolisado de cartilagem esternal de frango hidrolisado de baixo peso molecular, BioCell Collagen, na melhora dos sintomas relacionados à osteoartrite: um ensaio randomizado, duplo-cego e controlado por placebo. Jornal de Química Agrícola e Alimentar, Biblioteca Nacional de Medicina dos EUA, 25, abril de 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise, et al. A suplementação de peptídeo de colágeno em combinação com treinamento de resistência melhora a composição corporal e aumenta a força muscular em homens sarcopênicos idosos: um ensaio clínico randomizado O British Journal of Nutrition, Cambridge University Press, 28 de outubro de 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Ao informar as pessoas sobre como a Universidade Nacional de Ciências da Saúde fornece o conhecimento para as gerações futuras, a Universidade oferece uma ampla variedade de profissões médicas para medicina funcional.
Como você foi diagnosticado com doença celíaca, síndrome do intestino irritável, diverticulose / diverticulite ou síndrome do intestino permeável?
Arrotos excessivos, arrotos ou inchaço?
Distensão anormal após certos probióticos ou suplementos naturais?
Suspeita de má absorção nutricional?
Os problemas digestivos diminuem com o relaxamento?
Se você estiver enfrentando alguma dessas situações, poderá estar enfrentando problemas intestinais e talvez precise experimentar o Programa 4R.
Sensibilidades alimentares, artrite reumatóide e ansiedade têm sido associadas à permeabilidade gastrointestinal prejudicada. Essas várias condições podem ocorrer por vários fatores que podem afetar o trato digestivo. Se não tratada, pode potencialmente resultar da disfunção da barreira da permeabilidade intestinal, causando inflamação e condições graves de saúde que o intestino pode desenvolver. O programa 4R é usado para restaurar um intestino saudável no corpo e envolve quatro etapas. São eles: remover, substituir, reinocular e reparar.
Permeabilidade intestinal
A permeabilidade intestinal ajuda a proteger o corpo e garante que bactérias nocivas não entrem no intestino. Protege o corpo de potenciais fatores ambientais isso pode ser prejudicial e está entrando no trato digestivo. Pode ser toxina, microorganismos patogênicos e outros antígenos que podem prejudicar o trato digestivo, causando problemas. O revestimento intestinal é constituído por uma camada de células epiteliais que são separadas por junções estreitas. Em um intestino saudável, a junção estreita regula a permeabilidade intestinal, permitindo seletivamente que substâncias entrem e viajem através da barreira intestinal e impedindo a absorção de fatores prejudiciais.
Certos fatores ambientais podem danificar a junção estanque, e o resultado é que ela pode aumentar a permeabilidade intestinal, o que causa hiperpermeabilidade intestinal ou intestino permeável. Os fatores contribuintes podem aumentar a permeabilidade intestinal, como uma quantidade excessiva de gorduras saturadas e álcool, deficiências em nutrientes, estresse crônico e doenças infecciosas.
Com uma permeabilidade intestinal aumentada no intestino, ele pode permitir que os antígenos cruzem a mucosa intestinal e entrem na corrente sanguínea, causando uma resposta imune e uma inflamação no corpo. Existem certas condições gastrointestinais associadas à hiperpermeabilidade intestinal e, se não tratada, pode desencadear certas condições auto-imunes que podem causar danos ao organismo.
Programa 4Rs
O 4Rs é um programa que os profissionais de saúde aconselham seus pacientes a usar quando estão lidando com problemas digestivos disruptivos e ajudam a apoiar a recuperação intestinal.
Removendo o problema
O primeiro passo no programa 4Rs é remover patógenos nocivos e gatilhos de inflamação associados ao aumento da permeabilidade intestinal. Gatilhos como estresse e consumo crônico de álcool podem causar muitos danos ao corpo de um indivíduo. Portanto, o objetivo desses fatores nocivos do corpo é tratá-lo com medicamentos, antibióticos, suplementos e recomenda-se a remoção de alimentos inflamatórios da dieta, incluindo:
- Álcool
- glúten
- Aditivos alimentares
- Amidos e féculas
- certos ácidos gordos
- Certos alimentos aos quais uma pessoa é sensível
Substituindo os nutrientes
O segundo passo do programa 4Rs é substituir os nutrientes que estão causando os problemas intestinais através da inflamação. Certos nutrientes podem ajudar a reduzir a inflamação no intestino, assegurando que o trato digestivo esteja sendo apoiado. Existem alguns alimentos anti-inflamatórios que são nutritivos. Esses incluem:
- Alimentos ricos em fibras
- Omega-3s
- Azeite
- Cogumelos
- Ervas anti-inflamatórias
Existem certos suplementos que podem ser usados para apoiar a função digestiva, auxiliando e absorvendo os nutrientes para promover um intestino saudável. O que as enzimas digestivas fazem é ajudar a quebrar gorduras, proteínas e carboidratos no intestino. Isso ajudará a beneficiar indivíduos com comprometimento do trato digestivo, intolerância alimentar ou doença celíaca. Suplementos como suplementos de ácido biliar podem ajudar na absorção de nutrientes pela fusão de lipídios. Estudos declararam que os ácidos biliares foram usados para tratar o fígado, a vesícula biliar e o ducto biliar, evitando a formação de cálculos biliares após a cirurgia bariátrica.
Reinoculated The Gut
O terceiro passo é o programa 4rs para reinocular o micróbio intestinal com bactérias benéficas para promover uma função intestinal saudável. Estudos têm sido demonstrados que suplementos probióticos foram usados para melhorar o intestino, restaurando bactérias benéficas. Com esses suplementos, eles fornecem ao intestino um aprimoramento ao secretar substâncias anti-inflamatórias no corpo, ajudam a apoiar o sistema imunológico, alteram a composição microbiana do corpo e reduzem a permeabilidade intestinal no sistema intestinal.
Como probióticos são encontrados em alimentos fermentados e são considerados transitórios, pois não são persistentes no trato gastrointestinal e são benéficos. Surpreendentemente, eles ainda têm um impacto na saúde humana devido a influenciar o intestino, produzindo vitaminas e compostos antimicrobianos, proporcionando diversidade e função intestinal.
Reparando o intestino
A última etapa do programa 4Rs é reparar o intestino. Esta etapa envolve reparar o revestimento intestinal do intestino com nutrientes e ervas específicos. Essas ervas e suplementos podem ajudar a diminuir a permeabilidade intestinal e a inflamação no corpo. Algumas dessas ervas e suplementos incluem:
- Babosa
- Chios mástique
- DGL (alcaçuz desglicirrizinizado)
- A raiz do marshmallow
- L-glutamina
- Omega-3s
Polifenóis
- Vitamina D
- zinco
Conclusão
Uma vez que muitos fatores podem afetar adversamente o sistema digestivo de maneira prejudicial e podem contribuir para diversas condições de saúde. O principal objetivo do programa 4Rs é minimizar esses fatores que estão prejudicando o intestino e reduzindo a inflamação e o aumento da permeabilidade intestinal. Quando o paciente está sendo apresentado aos fatores benéficos que os 4Rs fornecem, ele pode levar a um intestino saudável e sarado. Alguns Produtos estão aqui para ajudar a apoiar o sistema gastrointestinal, apoiando os intestinos, melhorando o metabolismo do açúcar e visando os aminoácidos que se destinam a apoiar os intestinos.
O escopo de nossas informações limita-se a problemas de saúde quiroprática, músculo-esquelética e nervosa ou a artigos, tópicos e discussões sobre medicina funcional. Utilizamos protocolos funcionais de saúde para tratar lesões ou distúrbios do sistema músculo-esquelético. Nosso escritório fez uma tentativa razoável de fornecer citações de apoio e identificou o (s) estudo (s) de pesquisa relevante (s) que apóiam nossos posts. Também disponibilizamos cópias dos estudos de apoio à diretoria e / ou ao público, mediante solicitação. Para discutir melhor o assunto acima, não hesite em perguntar ao Dr. Alex Jimenez ou entre em contato em 915-850-0900.
Referências:
De Santis, Stefania, et al. Chaves nutricionais para modulação da barreira intestinal. Fronteiras na imunologia, Frontiers Media SA, 7 Dec. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ianiro, Gianluca, et al. Suplementação Enzimática Digestiva em Doenças Gastrointestinais. Metabolismo Atual de Medicamentos, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui, et al. Intestino Furado como um Sinal de Perigo para Doenças Autoimunes. Fronteiras, Frontiers, 5 May 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon, et al. Alimentos fermentados como fonte dietética de organismos vivos. Fronteiras em Microbiologia, Frontiers Media SA, 24, agosto de 2018, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R., et al. Rápida disrupção da função de barreira intestinal por gliadina envolve expressão alterada de proteínas juncionais apicais. Imprensa FEBS, John Wiley & Sons, Ltd, 8 de agosto de 2005, febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Sartor, R. Balfour. Manipulação terapêutica da microflora entérica em doenças inflamatórias intestinais: antibióticos, probióticos e prebióticos. Gastroenterologia, Biblioteca Nacional de Medicina dos EUA, maio 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
A dor crônica é um problema de saúde comum que afeta muitas pessoas nos Estados Unidos. Embora várias condições médicas, como fibromialgia e síndrome da dor miofascial, possam causar dor crônica, ela também pode se desenvolver devido a uma variedade de outros problemas de saúde. Estudos de pesquisa descobriram que a inflamação generalizada é a principal causa de dor crônica. A inflamação é um mecanismo de defesa natural a lesões, doenças ou infecções. Mas, se o processo inflamatório continuar por muito tempo, pode se tornar problemático.
A inflamação sinaliza ao sistema imunológico para curar e reparar o tecido danificado, bem como para se proteger contra bactérias e vírus. Como mencionado acima, no entanto, a inflamação crônica pode causar uma variedade de problemas de saúde, incluindo sintomas de dor crônica. Modificações saudáveis no estilo de vida podem ajudar a controlar a dor crônica, mas primeiro, vamos entender as causas comuns da dor crônica.
O que é inflamação aguda?
A inflamação aguda, por exemplo, ocorre após uma lesão ou algo tão simples como uma dor de garganta. É uma resposta natural com efeitos adversos, o que significa que funciona localmente na região onde o problema de saúde é encontrado. Os sinais comuns de inflamação aguda incluem inchaço, vermelhidão, calor, dor e perda de função, como afirma a National Library of Medicine. Quando a inflamação aguda se desenvolve, os vasos sangüíneos se dilatam, provocando o aumento do fluxo sanguíneo, e os glóbulos brancos da região lesada promovem a recuperação.
Durante a inflamação severa, compostos chamados citocinas são liberados pelo tecido danificado. As citocinas atuam como "sinais de emergência", que trazem as próprias células do sistema imunológico do corpo humano, bem como hormônios e inúmeros nutrientes para reparar o problema de saúde. Além disso, substâncias semelhantes a hormônios, conhecidas como prostaglandinas, causam a formação de coágulos sangüíneos para curar tecidos danificados, e estes também podem desencadear febre e dor como parte do procedimento inflamatório. Quando o dano ou lesão se recupera, a inflamação diminui.
O que é inflamação crônica?
Ao contrário da inflamação aguda, a inflamação crônica tem efeitos a longo prazo. A inflamação crônica, também conhecida como inflamação persistente, produz baixos níveis de inflamação em todo o corpo humano, como demonstrado pelo aumento dos marcadores do sistema imune localizados no sangue e nos tecidos celulares. A inflamação crônica também pode causar a progressão de várias doenças e condições. Níveis elevados de inflamação podem, às vezes, desencadear mesmo se não houver lesão, doença ou infecção, o que também pode fazer com que o sistema imunológico reaja.
Como resultado, o sistema imunológico do corpo humano pode começar a atacar células, tecidos ou órgãos saudáveis. Os pesquisadores ainda estão tentando entender as consequências da inflamação crônica no corpo humano e os mecanismos envolvidos nesse processo natural de defesa. Por exemplo, a inflamação crônica tem sido associada a uma variedade de problemas de saúde, como doenças cardíacas e derrame.
Uma teoria sugere que, quando a inflamação permanece nos vasos sanguíneos, ela pode estimular o acúmulo de placas. Segundo a American Heart Association, ou a AHA, se o sistema imunológico identifica a placa como um invasor externo, as células brancas do sangue podem tentar isolar a placa encontrada no sangue que flui pelas artérias. Isso pode criar um coágulo sanguíneo que pode bloquear o fluxo sanguíneo para o coração ou cérebro, fazendo com que ele se torne instável e se rompa. O câncer é outro problema de saúde associado à inflamação crônica. Além disso, de acordo com o Instituto Nacional do Câncer, danos no DNA também podem ser causados por inflamação crônica.
A inflamação persistente e de baixo grau freqüentemente não apresenta sintomas, mas os profissionais de saúde podem verificar se há uma proteína C-reativa, ou PCR, conhecida como ácido lipóico, um marcador de inflamação encontrado no sangue. Níveis elevados de PCR estão associados a um risco aumentado de doença cardiovascular. Níveis elevados de PCR podem ser encontrados em distúrbios crônicos como lúpus ou artrite reumatóide.
No caso de outras condições crônicas, como a fibromialgia, o sistema nervoso reage excessivamente à estimulação específica, no entanto, é a inflamação que causa sintomas de dor crônica. Subjetivamente, é quase impossível dizer a diferença entre a dor crônica causada por um sistema nervoso supersensível e a dor crônica causada pela inflamação generalizada. Além de procurar por pistas na corrente sanguínea, a nutrição de uma pessoa, hábitos de vida e exposições ambientais também podem promover inflamação crônica.
A inflamação é o mecanismo de defesa natural do sistema imunológico contra lesões, doenças ou infecções. Embora essa resposta inflamatória possa ajudar a curar e reparar tecidos, a inflamação crônica generalizada pode causar uma variedade de problemas de saúde, incluindo sintomas de dor crônica. Um equilíbrio a nutrição, incluindo uma variedade de dietas e jejum, pode ajudar a reduzir a inflamação. O jejum, também conhecido como restrição calórica, promove a apoptose celular e a recuperação mitocondrial. A dieta que imita o jejum, que faz parte do plano de dieta da longevidade, é um programa dietético que “engana” o corpo humano em um estado de jejum para experimentar os benefícios do jejum tradicional. Antes de seguir qualquer uma das dietas descritas neste artigo, consulte um médico.
Dr. Alex Jimenez DC, Insight CCST
Nutrição, Dietas, Jejum e Dor Crônica
Dietas anti-inflamatórias consistem principalmente de comer frutas e legumes frescos, peixes e gorduras. O plano de dieta mediterrânea, por exemplo, é uma dieta antiinflamatória que promove a ingestão de quantidades moderadas de nozes, a ingestão de muito pouca carne e a ingestão de vinho. As partes alimentares antiinflamatórias, como os ácidos graxos ômega-3, protegem o corpo humano contra o damago provocada pela inflamação.
Uma dieta anti-inflamatória também envolve ficar longe de alimentos que poderiam promover a inflamação. É ideal para diminuir a quantidade de alimentos que você come, que são ricos em gorduras trans e saturadas, como carnes. Além disso, uma dieta anti-inflamatória limita o consumo de carboidratos refinados e alimentos, como pão e arroz. Estes também promovem o corte na utilização de margarina e óleos que são embalados com ácidos graxos ômega-6, como girassol, cártamo e óleos de milho.
O jejum, ou restrição calórica, tem sido conhecido por diminuir o estresse oxidativo e retardar os mecanismos de envelhecimento em vários organismos. Os efeitos do jejum envolvem morte celular programada, ou apoptose, transcrição, eficiência energética móvel, biogênese mitocondrial, mecanismos antioxidantes e ritmo circadiano. O jejum também contribui para a autofagia mitocondrial, conhecida como mitofagia, onde os genes nas mitocôndrias são estimulados a sofrer apoptose, o que promove a recuperação mitocondrial.
O jejum intermitente pode ajudá-lo a combater a inflamação, melhorar a digestão e aumentar sua longevidade. O corpo humano foi projetado para sobreviver por longos períodos sem comida. Estudos de pesquisa demonstraram que o jejum intermitente pode ter mudanças positivas na composição geral da microbiota intestinal. Além disso, o jejum intermitente pode reduzir a resistência à insulina enquanto aumenta a resposta do sistema imunológico. Por fim, o jejum intermitente pode promover a produção de uma substância, conhecida como? -Hidroxibutirato, que bloqueia uma parte do sistema imunológico envolvida em doenças inflamatórias, além de reduzir substancialmente a produção de marcadores inflamatórios, como citocinas e a proteína C reativa , ou CRP, mencionado anteriormente.
O Plano de Dieta da Longevidade, apresentado no livro do Dr. Valter Longo, elimina o consumo de alimentos processados que podem causar inflamação, promovendo bem-estar e longevidade. Este programa dietético único, ao contrário da maioria das dietas tradicionais, não promove a perda de peso. Embora você possa experimentar redução de peso, a ênfase deste programa dietético exclusivo é a alimentação mais saudável. O plano de dieta da longevidade foi demonstrado para ajudar a ativar a renovação baseada em células-tronco, reduzir a gordura abdominal e prevenir perda óssea e muscular relacionada à idade, bem como desenvolver resistência ao desenvolvimento de doenças cardiovasculares, doença de Alzheimer, diabetes e câncer.
O jejum imitando a dieta, ou a febre aftosa, permite que você experimente os benefícios do jejum tradicional sem privar seu corpo de alimentos. A principal diferença da febre aftosa é que, em vez de eliminar completamente todos os alimentos por vários dias ou mesmo semanas, você só restringe sua ingestão de calorias por cinco dias fora do mês. A febre aftosa pode ser praticada uma vez por mês para ajudar a promover a saúde e bem-estar geral.
Enquanto qualquer um pode seguir a febre aftosa por conta própria, o ProLon O jejum que imita a dieta oferece um programa de refeição de dia 5 que foi embalado individualmente e rotulado para cada dia, que serve os alimentos que você precisa para a febre aftosa em quantidades e combinações precisas. O programa de refeição é composto de alimentos à base de plantas prontos para comer ou fáceis de preparar, incluindo barras, sopas, lanches, suplementos, um concentrado de bebida e chás. Antes de iniciar o ProLon jejum imitando dieta, programa de refeições de 5 diasou qualquer uma das modificações de estilo de vida descritas acima, por favor, certifique-se de conversar com um profissional de saúde para descobrir qual tratamento de dor crônica é ideal para você.
O escopo de nossas informações é limitado a quiropraxia, questões de saúde da coluna vertebral e artigos sobre medicina funcional, tópicos e discussões. Para discutir mais sobre o assunto acima, sinta-se à vontade para perguntar ao Dr. Alex Jimenez ou entrar em contato conosco. 915-850-0900 .
Curated pelo Dr. Alex Jimenez
Discussão de tópico adicional: Dor aguda nas costas
Dor nas costas é uma das causas mais prevalentes de incapacidade e perdeu dias de trabalho em todo o mundo. A dor nas costas atribui-se à segunda razão mais comum para visitas a consultórios, superada apenas por infecções respiratórias superiores. Aproximadamente 80 por cento da população experimentará dor nas costas pelo menos uma vez ao longo da vida. Sua espinha é uma estrutura complexa composta de ossos, articulações, ligamentos e músculos, entre outros tecidos moles. Lesões e / ou condições agravadas, como hérnia de discos, pode eventualmente levar a sintomas de dor nas costas. Lesões esportivas ou acidentes automobilísticos geralmente são a causa mais frequente de dor nas costas, no entanto, às vezes, o mais simples dos movimentos pode ter resultados dolorosos. Felizmente, opções alternativas de tratamento, como quiropraxia, podem ajudar a aliviar a dor nas costas através do uso de ajustes espinhais e manipulações manuais, melhorando o alívio da dor.
XYMOGEN s Fórmulas profissionais exclusivas estão disponíveis através de profissionais de saúde licenciados selecionados. A venda pela internet e o desconto de fórmulas XYMOGEN são estritamente proibidos.
Orgulhosamente, Dr. Alexander Jimenez faz com que as fórmulas XYMOGEN estejam disponíveis apenas para pacientes sob nossos cuidados.
Por favor, ligue para o nosso escritório para que possamos atribuir uma consulta médica para acesso imediato.
Se você é um paciente de Clínica médica e de Quiropraxia para lesões, você pode perguntar sobre o XYMOGEN chamando 915-850-0900.
Para sua conveniência e revisão do XYMOGEN produtos por favor reveja o seguinte link. *Catálogo XYMOGENBaixar
* Todas as políticas XYMOGEN acima permanecem estritamente em vigor.
Aderir a uma dieta específica para manter uma nutrição adequada pode, às vezes, tornar a alimentação estressante. Modificações naturais no estilo de vida são a chave para mudar seus hábitos alimentares e isso pode ajudá-lo a viver uma vida mais longa e saudável. O plano de dieta da longevidade, criado pelo Dr. Valter Longo, é uma seleção de orientações práticas de alimentação que se concentra em mudar seus padrões alimentares para alcançar saúde e bem-estar geral.
As regras do plano de dieta da longevidade
Por apenas seguindo as dicas nutricionais abaixo, você pode revisar seu plano de dieta atual e começar a comer mais saudável sem todo o stress de uma dieta tradicional. O plano de dieta da longevidade elimina o consumo de alimentos processados que podem causar uma variedade de problemas de saúde e estimula o consumo de nutrientes que promovem a longevidade. Este programa dietético exclusivo compartilha os resultados de aproximadamente 25 anos de estudos de pesquisa, tudo em uma solução simples que pode ajudar as pessoas a experimentar bem-estar geral através de uma nutrição adequada.
No entanto, ao contrário da maioria das dietas tradicionais, o Plano de Dieta da Longevidade não promove a perda de peso. Embora você possa experimentar redução de peso, a ênfase deste programa dietético exclusivo é uma alimentação mais saudável. O Plano de Dieta da Longevidade demonstrou ajudar a ativar a renovação baseada em células-tronco, perder peso e reduzir a gordura abdominal, prevenir a perda óssea e muscular relacionada à idade, construir resistência ao desenvolvimento de doenças cardiovasculares, doença de Alzheimer, diabetes e câncer, também como estender a longevidade. Abaixo, vamos resumir as 8 dicas nutricionais mais comuns do Plano de Dieta da Longevidade que podem ajudar a tornar sua vida mais longa e saudável.
O plano de dieta da longevidade é um programa de dieta exclusivo projetado pelo Dr. Valter Longo para promover a saúde geral, bem-estar e longevidade. Através de modificações simples no estilo de vida, as pessoas podem mudar seus hábitos alimentares e tirar proveito dos muitos benefícios para a saúde deste programa dietético. Seguindo uma dieta de pescatarian e seguindo o Dieta de imitação de jejum ProLon Entre as outras dicas nutricionais descritas abaixo, as pessoas podem viver vidas mais longas e saudáveis. Dietas tradicionais muitas vezes podem ser difíceis e estressantes para seguir, no entanto, o plano de dieta de longevidade é um programa de dieta prático e exclusivo que pode ser adequado para muitas pessoas.
Dr. Alex Jimenez DC, Insight CCST
8 Dicas Nutricionais do Plano de Dieta de Longevidade
Siga uma dieta Pescatariana
Como parte do plano de dieta da longevidade, siga uma dieta de pescatarian, que é quase 100 por cento de plantas e peixes. Além disso, certifique-se de limitar o consumo de peixe a duas ou três porções por semana, evitando peixes com maior teor de mercúrio, como atum, peixe-espada, cavala e alabote. Se você estiver com 65 e começar a sentir redução de massa muscular, força e gordura, acrescente mais peixes à sua dieta junto com outros alimentos de origem animal, incluindo ovos e queijos específicos, como queijo feta ou pecorino, e iogurte de cabra. leite.
Não coma muita proteína
De acordo com o plano de dieta da longevidade, devemos comer 0.31 0.36 gramas de proteína por quilo de gordura corporal todos os dias. Se você pesa 130lbs, você deve comer cerca de 40 para 47 gramas de proteína por dia, ou um equivalente de filés 1.5 de salmão, 1 xícaras de grão de bico ou xícaras de lentilhas 2 1 / 2, das quais 30 gramas devem ser consumidas em uma refeição. Se você pesa 200 em 220lbs, deve ingerir cerca de 60 em 70 gramas de proteína por dia, ou o equivalente a dois filés de salmão, 3 1 / 2 xícaras de lentilhas ou xNUMX 1 / 1 xícaras de grão-de-bico. O consumo de proteínas deve ser aumentado após a idade 2. Para a maioria de nós, um aumento de 65 para 10 por cento, ou 20 para 5 gramas a cada dia, é o suficiente. Finalmente, a Dieta da Longevidade é livre de proteínas animais, como carne vermelha, carne branca e aves, com exceção de proteínas animais em peixes. Este programa dietético único é comparativamente alto em proteínas vegetais como legumes e nozes para otimizar a saúde e bem-estar.
Aumente as gorduras boas e carboidratos complexos
Como parte do plano de dieta da longevidade, você deve ingerir maiores quantidades de gorduras poliinsaturadas, como as encontradas em salmão, amêndoas, nozes e azeite, enquanto você deve ingerir quantidades menores de gorduras saturadas, hidrogenadas e trans. Da mesma forma, como parte do Plano de Dieta da Longevidade, você também deve ingerir carboidratos complexos, como os encontrados no pão de trigo integral, legumes e verduras. Certifique-se de limitar a ingestão de massa, arroz, pão, frutas e sucos de frutas, que podem ser convertidos em açúcares quando chegarem ao seu intestino.
Tome suplementos dietéticos
O corpo humano precisa de proteínas, ácidos graxos essenciais como ômega-3 e ômega-6, vitaminas, minerais e até mesmo açúcares para funcionar corretamente. Sempre que sua ingestão de certos nutrientes se torna muito baixa, os métodos de reparo, substituição e defesa do corpo humano podem diminuir ou parar, permitindo que fungos, bactérias e vírus causem danos que podem levar a uma variedade de problemas de saúde. Tome suplementos vitamínicos e minerais, especialmente para ômega-3, conforme recomendado pelo seu profissional de saúde.
Coma vários alimentos do seu Ancestry
Para ingerir todos os nutrientes necessários, você precisa ingerir uma ampla variedade de alimentos, mas é melhor escolher alimentos comuns na mesa de seus pais, avós e bisavós. Por exemplo, em muitos países do norte da Europa onde o leite é geralmente consumido, a intolerância à lactose é relativamente rara, enquanto a intolerância à lactose é bastante comum nos países do sul da Europa e da Ásia, onde o leite não fazia parte da dieta convencional dos adultos. Se uma pessoa de ascendência japonesa residente nos Estados Unidos decidir de repente começar a beber leite, o que provavelmente foi raramente servido na mesa de jantar de seus avós, eles provavelmente começarão a se sentir mal. Os problemas mais comuns nesses casos são intolerâncias ou autoimunidades, como a resposta a alimentos ricos em glúten, como pão e massa, vistos em pessoas com doença celíaca. Embora mais evidências sejam necessárias, é possível que as intolerâncias alimentares possam estar relacionadas a muitos distúrbios autoimunes, incluindo diabetes, colite e doença de Crohn.
Coma duas refeições por dia e um lanche
De acordo com o Plano de Dieta da Longevidade, é ideal tomar o café da manhã e uma refeição importante, além de um lanche nutritivo de baixas calorias e baixo teor de açúcar todos os dias. Enquanto para algumas pessoas pode ser recomendado comer três refeições e um lanche todos os dias. Muitas diretrizes nutricionais recomendam que devemos comer cinco a seis refeições todos os dias. Quando as pessoas são aconselhadas a comer com frequência, muitas vezes pode ser difícil para elas regular a ingestão de calorias. Nos últimos 70 anos, aproximadamente 100% da população dos Estados Unidos foi considerada com sobrepeso ou obesidade. É muito mais difícil comer demais no Plano de Dieta da Longevidade se você comer apenas duas refeições e meia por dia. Seriam necessárias grandes porções de legumes, vegetais e peixes para atingir a quantidade que levaria ao ganho de peso. A alta nutrição das refeições, mais a quantidade da refeição, envia um sinal ao estômago e ao cérebro de que você comeu o suficiente. Este único sistema de refeição principal pode às vezes ter que ser dividido em duas refeições para evitar problemas de digestão. Adultos e pessoas mais velhas com tendência à perda de peso devem comer três refeições por dia. Para pessoas que estão tentando perder peso, bem como para pessoas com sobrepeso ou obesas, o melhor conselho nutricional seria tomar o café da manhã diariamente; jantar ou almoçar, mas não ambos, e substituir a refeição perdida por um lanche contendo menos de 3 calorias e não mais do que 5 a XNUMX g de açúcar. A refeição que você pula depende do seu estilo de vida, no entanto, não é recomendável pular o café da manhã devido a seus problemas adversos à saúde. O benefício de pular o almoço é mais tempo livre e energia. No entanto, há uma desvantagem em comer um jantar farto, especialmente para pessoas que sofrem de refluxo ácido ou problemas de sono. A desvantagem de pular o jantar, no entanto, é que isso pode eliminar a refeição social do dia.
Coma dentro de uma janela 12-Hour todos os dias
Outro hábito alimentar comum adotado por muitos centenários é a restrição do tempo de comer ou limitar todas as refeições e lanches em uma janela de 12-hora todos os dias. A eficiência deste método foi demonstrada em estudos de pesquisa em humanos e animais. Geralmente, você toma café da manhã no 8 am e depois janta por 8 pm. Uma janela de dez horas ou menos pode ser ainda melhor para perda de peso, mas é consideravelmente mais difícil de manter e pode aumentar o risco de desenvolver efeitos colaterais, como cálculos biliares e até mesmo aumentar potencialmente a chance de desenvolver doença cardiovascular. Você não deve comer de três a quatro horas antes de dormir.
Siga a dieta de imitação de jejum ProLon
Pessoas saudáveis com idade inferior a 65 devem seguir o Dieta de imitação de jejum ProLon , programa de refeições de 5 dias pelo menos duas vezes por ano. A febre aftosa é um dos princípios fundamentais promovidos pelo plano de dieta da longevidade. O jejum dieta imitando oferece os mesmos benefícios de saúde do jejum, sem realmente jejum. Ao comer 800 em 1,100 calorias em quantidades precisas e combinações de alimentos que foram embalados individualmente e rotulados para cada dia, você pode "enganar" o corpo humano em um estado de jejum. Através de vários estudos de pesquisa, o Dr. Valter Longo descobriu que, ao privar o corpo de alimentos dessa maneira, nossas células começam a quebrar e regenerar nossos tecidos internos, através de um processo conhecido como autofagia, matando e substituindo ou regenerando células danificadas. Além disso, o jejum pode reverter vários problemas de saúde, destruir as células cancerígenas e reduzir significativamente a possibilidade de desenvolver a doença de Alzheimer.
Com o Plano de Dieta da Longevidade apresentado no livro do Dr. Valter Longo, você comerá melhor, se sentirá melhor e, embora não seja um plano de emagrecimento, pode até perder alguns quilos. Você não terá que considerar regras alimentares complexas e fazer escolhas difíceis com este programa dietético exclusivo. Depois de pegar o jeito dessas modificações no estilo de vida, você será capaz de melhorar sua saúde geral e bem-estar, bem como os longevidade. A O escopo de nossa informação é limitado à Quiropraxia, questões de saúde da coluna vertebral e tópicos de medicina funcional. Para discutir mais sobre o assunto, sinta-se à vontade para perguntar ao Dr. Alex Jimenez ou contate-nos 915-850-0900 .
Curated pelo Dr. Alex Jimenez
Discussão de tópico adicional: Dor aguda nas costas
Dor nas costas é uma das causas mais prevalentes de incapacidade e perdeu dias de trabalho em todo o mundo. A dor nas costas atribui-se à segunda razão mais comum para visitas a consultórios, superada apenas por infecções respiratórias superiores. Aproximadamente 80 por cento da população experimentará dor nas costas pelo menos uma vez ao longo da vida. Sua espinha é uma estrutura complexa composta de ossos, articulações, ligamentos e músculos, entre outros tecidos moles. Lesões e / ou condições agravadas, como hérnia de discos, pode eventualmente levar a sintomas de dor nas costas. Lesões esportivas ou acidentes automobilísticos geralmente são a causa mais frequente de dor nas costas, no entanto, às vezes, o mais simples dos movimentos pode ter resultados dolorosos. Felizmente, opções alternativas de tratamento, como quiropraxia, podem ajudar a aliviar a dor nas costas através do uso de ajustes espinhais e manipulações manuais, melhorando o alívio da dor.
XYMOGEN s Fórmulas profissionais exclusivas estão disponíveis através de profissionais de saúde licenciados selecionados. A venda pela internet e o desconto de fórmulas XYMOGEN são estritamente proibidos.
Orgulhosamente, Dr. Alexander Jimenez faz com que as fórmulas XYMOGEN estejam disponíveis apenas para pacientes sob nossos cuidados.
Por favor, ligue para o nosso escritório para que possamos atribuir uma consulta médica para acesso imediato.
Se você é um paciente de Clínica médica e de Quiropraxia para lesões, você pode perguntar sobre o XYMOGEN chamando 915-850-0900.
Para sua conveniência e revisão do XYMOGEN produtos por favor reveja o seguinte link. *Catálogo XYMOGENBaixar
* Todas as políticas XYMOGEN acima permanecem estritamente em vigor.
A ferramenta Find A Practitioner do IFM é a maior rede de referência em Medicina Funcional, criada para ajudar os pacientes a localizar profissionais de Medicina Funcional em qualquer lugar do mundo. Os Praticantes Certificados IFM são listados em primeiro lugar nos resultados da pesquisa, devido à sua extensa educação em Medicina Funcional